Genitalia of sexual reproductive species, especially intromittent organs in copulation, are morphologically complicate and biologically important organs that are attracting many biologists. Beetle genitalia are the organ of copulation and join directly sexual reproduction, more exactly, the organ to fulfil intromission and sperm transportation so that to get a successful insemination. Genital morphological variations are from one side to be used to distinguish different species (morphological or taxonomic species) and, on the other, to play a role in reproductive isolation between species when the species reproduce sexually. Genital morphology and its variations bridge essentially both the morphological and the biological species concepts; this is especially true if we consider again and more closely the classical but reactivated hypothesis of lock-and-key. It was generally recognized that the evolution of genitalia and attached membranous structure, particularly the copulary organ, was promoted by sexual selection and/or sexual conflict.
The primary aim of the present study is to investigate the morphology and variations of the endophallus in the phylogenetic context, using the phylogenetically enigmatic staphylinid tribe Xantholinini as a case study. To optimize the documentation of the everted endophalli a novel technique of micro-operating live males was developed. We observed five different types in the five examined species (Fig. 1; Fig. 3 as a representative of Type I). The studied genera of the tribe were recovered monophyletic with strong support, and the relationships were identical with both methods with strong or moderate support. The support values were distinctly stronger for the molecular data alone, as compared to analyses based on the combined matrix (Fig. 3). To assess correlations between modifications of the sclerotized part of the median lobe and the endophallus on a broader scale, especially gradual reduction of the former and increasing complexity of the latter, we selected 99 genera (more than 80% of the total) and 456 species of Xantholinini. We found that the length of the median lobe was negatively correlated with the endophallus length (r=0.31, p<0.01; Fig. 4A). Moreover, the length of the median lobe is negatively correlated with the structural complexity of the endophallus, with an increasingly complicated arrangement of sclerotized spines and membranous elements (r=0.45, p<0.01; Fig. 4B).
Our investigation of the male genitalia of Xantholinini revealed conspicuous evolutionary innovations of a clearly understudied part of the intromittent organ, the membranous endophallus. Our phylogenetic analysis shows that endophallus modifications and reduction of the rigid aedeagus (intromittent organ or penis) and parameres are evolutionary innovations of certain subunits of the tribe.
The first author Dr. Yu-Lingzi Zhou, has begun this study in her PhD study and continued it during her three year of Postdoc in the laboratory of Prof. Hongzhang Zhou, Institute of Zoology, Chinese Academy of Sciences. Afterwards, she got a grand from the Chinese government special supporting her to study overseas so that she can conduct her researches in the labor of Prof. Adam Slipiński from Australian National Insect Collection, CSIRO (Australia) and collaborate with Prof. Rolf G. Beutel from Friedrich-Schiller-Universitat Jena (Germany). This study was published on Zoological Journal of the Linnean Society. This project was also supported by the National Natural Science Foundation of China Grants No. 31402008, and International Postdoctoral Exchange Fellowship No. 20150064.
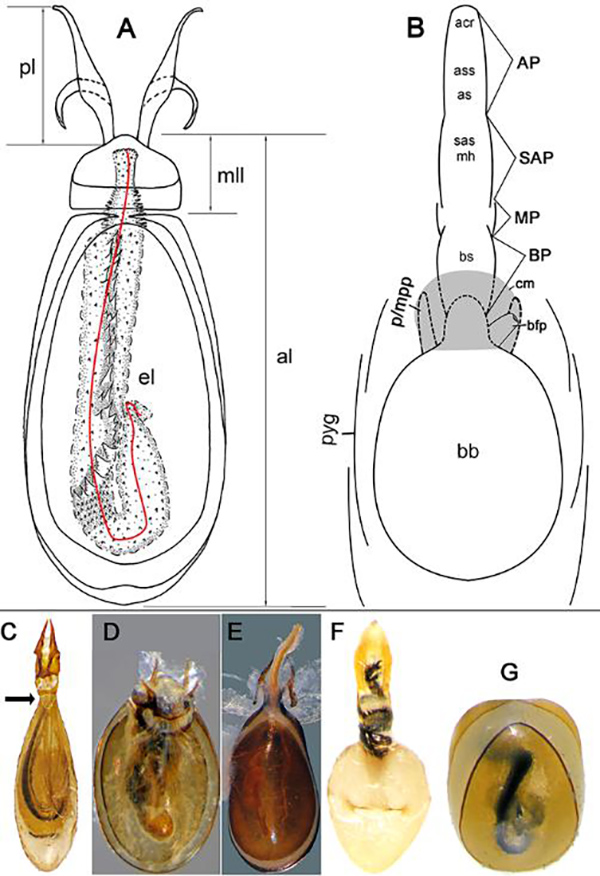
Figure 1. Schematic drawings of the fully eversible endophalli of four different types. Type I: Thyreocephalus hongkongensis (Redtenbacher, 1867); Type II: Xanthophius filum (Kraatz, 1859); Type IV: Megalinus flavus (Bordoni, 2002); Type V: Megalinus japonicus (Sharp, 1874).
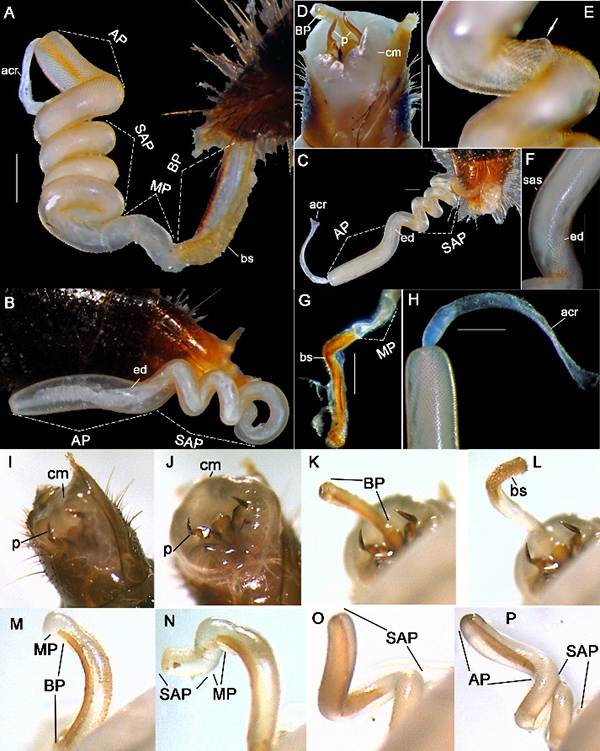
Figure 2. Endophallus Type I of Thyreocephalus hongkongensis (Redtenbacher, 1867). A) Fully everted (BP, MP, SAP, AP), ventrolateral view. B–C) SAP, AP of partly everted endophallus, dorsolateral view. D) Dilated cm. E) Junction point of acr (white arrow). F) sas and ejaculatory duct (ed). G) Dissected bs and MP. H) acr. I–J) cm and p everted. K–L) Phase one: BP everted. M) Phase two: MP everted. N–O) Phase three: SAP everted. P) Phase four: AP everted. acr: apical connective ribbon; bs: basal spines; cm: connective membrane; p: parameres; sas: sub-apical spines. Scale bars 0.30 mm.
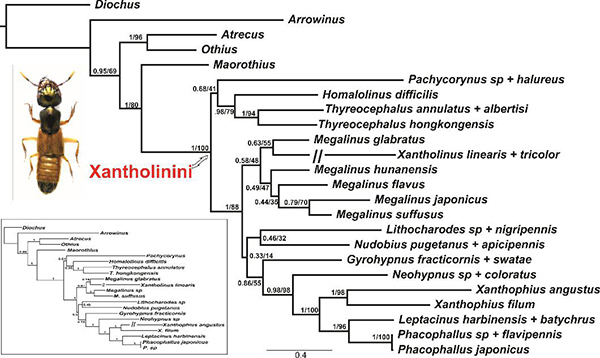
Figure 3. Bayesian phylogenetic tree of Xantholinini, based on the combined dataset including three genes: COI, 28S, wingless, and 26 morphological characters. Posterior probability and Bootstrap values are shown for each clade. Consistent phylogenetic tree obtained with BI and ML with molecular data only shown in blue rectangle.
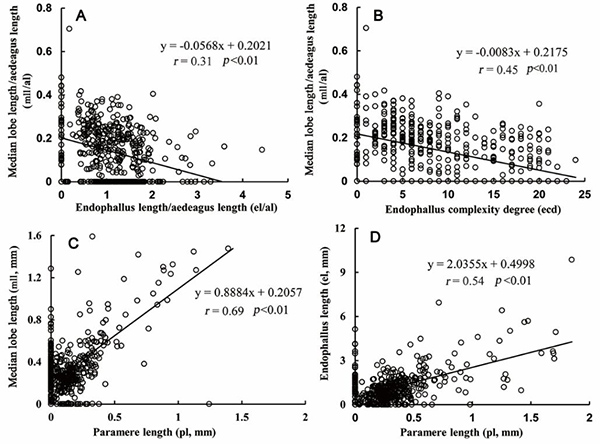
Figure 4. Correlations between median lobe, paramere and endophallus. A) Median lobe length / aedeagus length (mll/al) vs. endophallus length / aedeagus length (el/al). B) Median lobe length / aedeagus length (mll/al) vs. endophallus complexity degree (ecd). C) Median lobe length (mll) vs. paramere length (pl). D) Endophallus length (el) vs. paramere length (pl). pl: paramere length; mll: median lobe length; al: aedeagus length; el: endophallus length. Line represents conventional linear regression.
